第20回 「霊長類の社会構造」
マハレでの餌付け成功後、伊谷先生は生態人類学への傾斜を深め、霊長類のフィールド・ワークの最前線から身を退き始めます。これは他の研究者の皆さんはもとより*、教え子の方々にも予想外だったようですが、例えば、マハレでは、基地の運営は西田利貞さんに任せ、自らは10年近く付き合っていたトングウェを対象とした生態人類学を始めます。(*:1970年代半ば、研究会等で動物生態学の小野勇一さんや心理学の岡野恒也さんから、まだ修士に入ったばかりの私に向かって「伊谷さんはもうサルをやらないのか?」とかなり真剣に尋ねられたことを覚えています)

左2枚は1966年頃の初期のカソゲの餌場。右2枚は1975年頃のカソゲ・カンシアナ基地
この転進の動機について、西田さんは先生への追悼文「遙かなるイプンバの想い出」(2002)で「第1は1960年の第2回アフリカ調査のとき狩猟採集民ピグミーに会われたことであろう。第2に、京大に霊長類研究所ができ、そこで本格的にサルの研究をやれるのだから、自然人類学講座ではヒトをやらなければならない、と考えられたこと。しかも、池田先生が人類学の研究室なんだからヒトの研究をやるべきだと、しきりにおっしゃっていた。第3に、サルの研究だけでは学生が食えなかろう、ということだ」とします。
しかしながら、第8回で触れた1954年の高崎山での水原洋城さんへの発言や、第12回で紹介したハッザについての「自分の手でこの人々の生活を明らかにしえないのがいかにもつらい」等の述懐をふり返れば、そもそも研究生活の始まりから、個人的な深い思い入れがあったと受け取るべきかもしれません。

意識的にトングウェの生活を記録し始めた1970〜71年の写真。しかし、残された時間は短く、1970年代半ばのタンザニア政府によるウジャマー村(集村化)計画で、原野での生活は霧散します
一方で、フィールドの最前線から退いても、霊長類学から離れたわけではありません。この点についても、西田さんは別の追悼文で『高崎山のサル』での今西・伊谷両氏のやりとりをとりあげ、伊谷先生の「今西さんは、やりたい野外調査をわざとやらず、学生にやらせたのだと思う。自分は理論をやるべきや、と考えはったんやろ」という言葉を紹介し、それを自分でも踏襲する=観察は若い世代にまかせ、彼らを理論家として支えようとしたのではないのか、と推測しています。
いずれにしても1970年代以降、伊谷先生は扱う分野を意図的に二つに絞ります。生態人類学でのExtensiveなフィールド・ワークと、霊長類の社会構造とその進化の理論化です。同時に、研究室ならびに研究体制を整えていきます。
(1)アフリカ等でのフィールド運営を確立された方には、その地・分野での采配をまかす(故原子令三さんの表現を借りると“一国一城の主”です)。
(2)学部では講義で霊長類学・人類学に関心を持つ学生を惹きつけ、ゼミや卒論指導を通じて自然人類や霊長研(後にはアフリカセンターも)等へ導く。
(3)自然人類/人類進化論は、修士課程は主として国内研究を指導するが、博士課程は国外のフィールドで、上記の一国一城の主等に預ける。そこで院生が得た観察・知識を帰国後のゼミで聞き、そのエッセンスを咀嚼・消化し、院生を指導するとともに、自らの理論を練り直す(=現場の報告からのフィードバック)。そこでは伊谷さんは“聞き役”に重きをおいていたような印象があります。自分がその場にいなかったフィールドでの観察を理解する=観察者/報告者の視点にアイデンティファイするため、とりあえず“聞く”ことに専念する。それは、もちろん、タイパ・コスパ流行りの昨今では、非効率で時代遅れと受け取る方も多いかもしれませんが。
ところで、ゼミでの発表者は、動物学教室にとどまらず、霊長類研究所はもとより、農学部や文学部等他学部の出身者も珍しくはありませんでした。その結果、今西さんとはまた違う形での研究ネットワークが広がります。これも“伊谷スクール”としての新たな展開となります。霊長類研究所出身で、ブッシュマンの研究者の菅原和孝さんは“若い世代へのメッセージ”に「伊谷さんは卓越したフィールドワーカーであると同時に、若者の教育に全身全霊を投入された。直接の弟子でもない私が持ちこんだ原稿を、徹底的に添削指導されたばかりか、原稿用紙数枚にわたって厳しいコメントをつけてくださったことを一生忘れない。伊谷さんは根本のところでアナーキストであり、自然哲学者である。西欧の理論の受け売りをやめて、自分がほんとうに面白いと思うことを、妥協せずに、忍耐づよく、一生追究してください」と語っています。研究と研究室の世代交代をも意識した、見事な戦略であった、と言うべきかもしれません。
さて、1970年前後、世界に生息する多様な霊長類についての知識が蓄積され始められると、伊谷先生は霊長類学の社会構造の進化について系統立てて考察し得る時期になったと判断します。それは同時に今西さんとともに1948年以来追求してきた、ヒトの社会の起原を考えることにもつながるはずでした。その時、伊谷先生が基盤としたのは「共通祖先から派生したものは似ているはずだ」という分類・系統学のパラダイムと思われます。それは今西さんの初期の盟友だった岩田久二雄さんや、その後坂上昭一さんらが昆虫を対象に展開した比較習性学がモデルだったかもしれません(下図)。
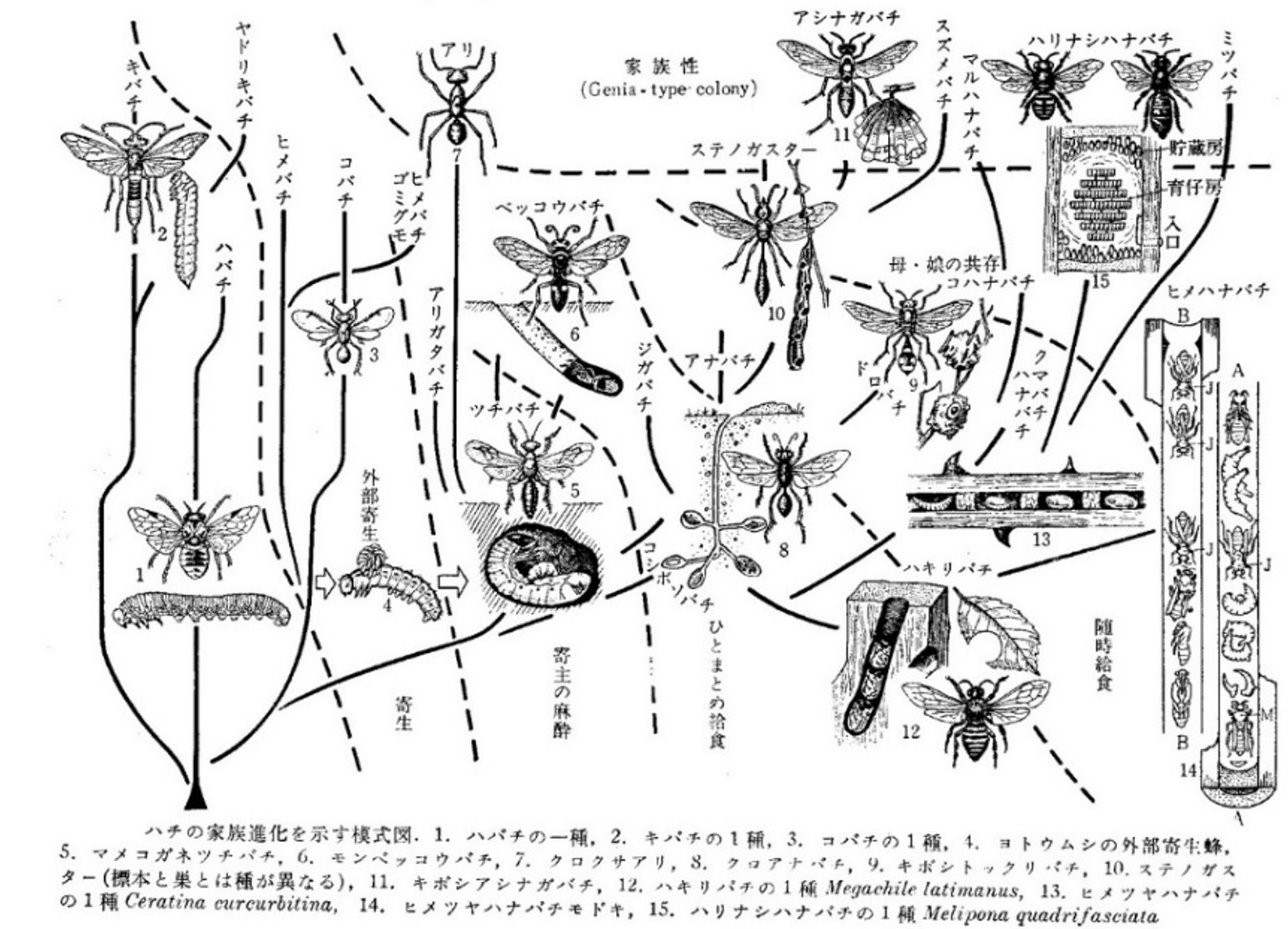
ハチの社会性進化の模式図:『比較生態学』(伊藤嘉昭、1966)から引用
その最初の試みが、1972年出版の『霊長類の社会構造』です。そこでは、伊谷先生は、通時的な視点で、集団が存在しない単独性の「要素的社会」から、「霊長類の社会は構造を異にする二つの系統(ペア型と群れ型)が進化した」と見立てます。
当時、ニホンザルでは、第5回で紹介した『高崎山のサル』掲載の図(いわゆる二重同心円モデル)が含意する“閉鎖的社会構造”が、その後の高宕山・嵐山等での長期データで否定され、オスは性成熟前後に出自群を離脱、他群に移籍することが一般的なことが明らかにされていました。伊谷先生はこれを、母系的な社会構造ととらえ直します。
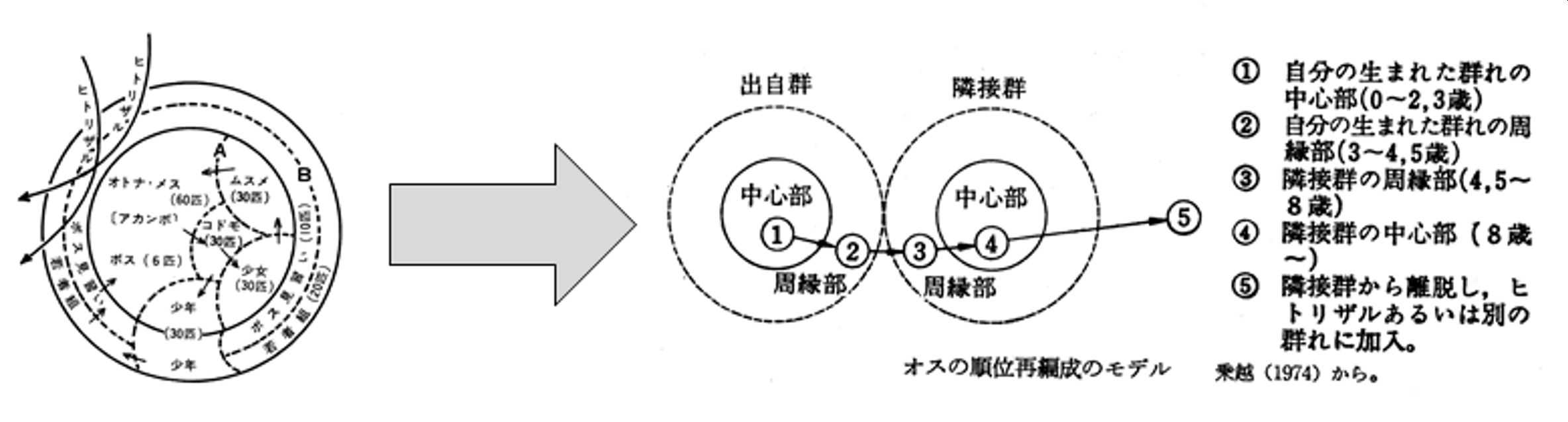
『高崎山のサル』の閉鎖的モデルから、嵐山群での半閉鎖的モデルへ
こうして、『高崎山のサル』で提示した(そして、いつの間にか定説・神話化してしまった)共時的視点からの“閉鎖的なBSU”のイメージを拭い去り、“通時的”な視点から“世代を超えての集団の継承”という基準で見直すというパラダイム・シフトを演じます。(その点に関連して、ニホンザルを含めてオナガザル科等の群れをたんに“female-bonded”と形容するか[Wrangham, 1980]、それとも“matrilineal”と表現する[Imanishi, 1960、Itani, 1977]かは、結構、重要なポイントかもしれません)
いずれにしても、『霊長類の社会構造』ではこの独自の基準で、霊長類の社会を以下のように大胆に切り分けました。
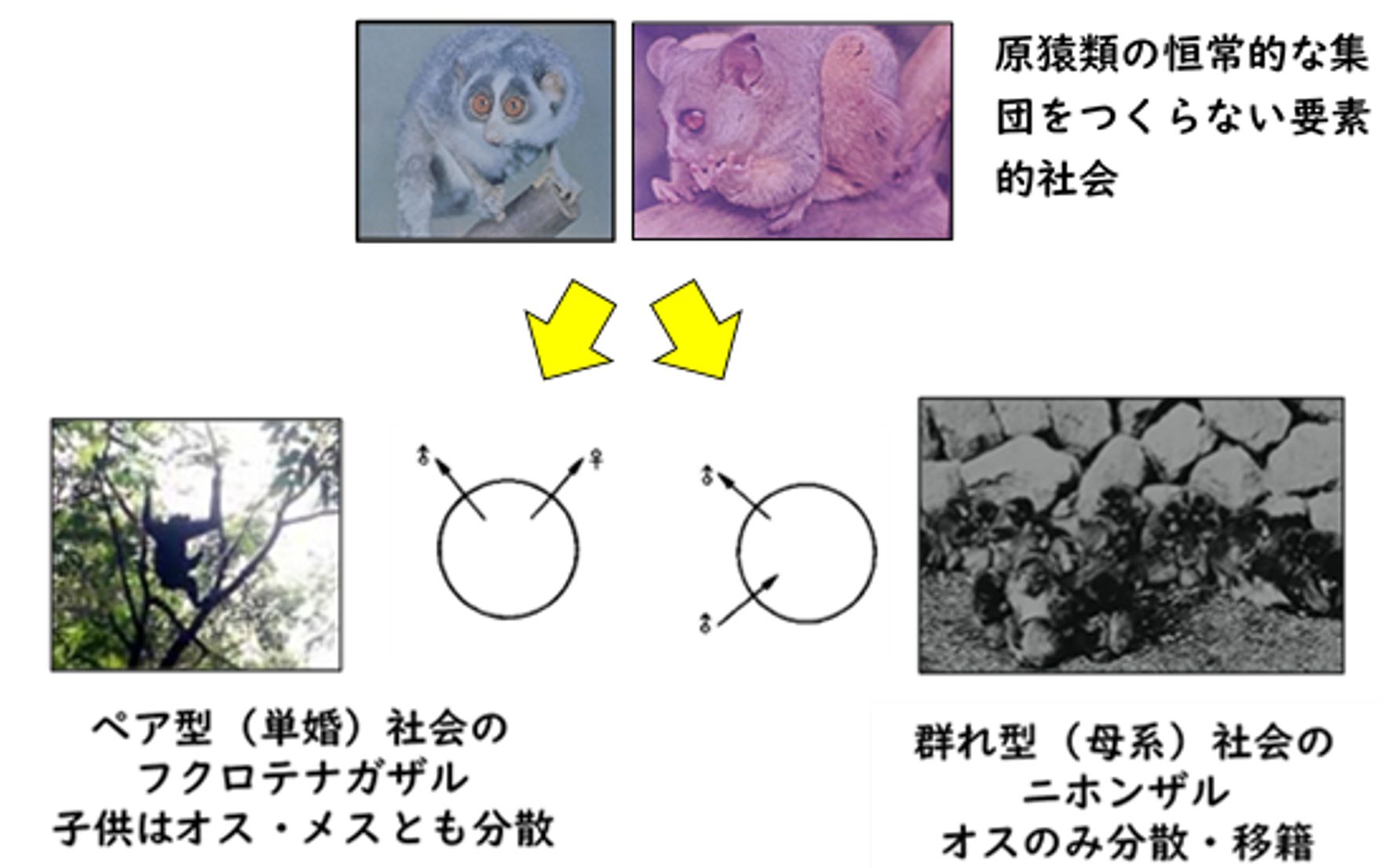
『霊長類の社会構造』での解釈
まず、ペア型とは、「コドモはオスもメスも出自集団から離れる」ことが特徴で、その典型がテナガザルです。どちらの性のコドモも出自群にとどまらないため、この集団は1世代限りで、集団の世代を超えた継承は考えられません。
一方、群れ型とは「オスのコドモだけが出自群を離れ、他群に移籍する=母系的関係で結びついたメスが群れを継承していく」構造です。ペア型との違いは「メスが出自群を離れるか、それともメスが同性との共存を許容するか」という点にあるとした上で、どうやら系統群(例えば「科」)ごとにどのタイプに属するか決まっているようだと考えます。こうして、ニホンザルを含むオナガザル科の社会構造は群れ型であるのに対して、類人猿はペア型ではないのか、と推論しました。この思考過程において、霊長類の社会を論じる上で、“生態”よりも“系統”を重視する姿勢が強まりますが、その点はまた改めて取り上げたいと思います。

対照的なニホンザルのオスとメスの生活史。左は「ヒトリザル“ミミナシ”。交尾期になると高崎山の群れに接近、リーダーのジュピターと激しく対し、相互に示威を繰り返した」。右は「高崎山での雌の子供への毛づくろい」。オスとは対照的に、メスは生まれた群れにとどまり、娘、孫娘等の母系で順位等を継承します。
ところで、この間、とくにニホンザルの母系的構造について、恩師今西さんとの間で意見の食い違いが目立ち始めます。今西さんはこうした考え方を、人類学/社会学から生物学への“逆行”(=生物学主義)ととらえ、忌避すべきものとして批判を強めます。その姿勢の背景には、今西さんなりの“理論”がありました。今西さんはニホンザル研究の初期、オスのコドモが成長する際、ボスザル/リーダーにアイデンティファイ(同一視/同一化)することで「群れ本位の行動」を体得すると考えていました。いわゆるアイデンテイフィケーション・セオリーです。今西さんの立場からすれば、「オスの群れ落ち」という報告自体が自説に影を落とすように感じられました。せっかく同一視で群れ本位の行動を身に付けたはずのオスが、どうして群れを捨て、「ヒトリザルの道を選んだということが、私には腑に落ちかねる」と漏らします。伊谷先生がさらにニホンザルの群れを母系的構造としてとらえるようになると、今西さんの戸惑いはさらに広がったわけです。(いまの視点でこの論争を振り返れば、“理論の奴隷”を厭ったはずの今西さんが、現場から離れ、かつての理論に固執し、新しい発見を消化できなくなったという、“理論家”の悲哀が感じられます)
なお、これは後知恵に過ぎませんが、この時、アイデンテイフィケーションのイメージを“ボスザル”ではなく、 “娘が母親のすぐ下の順位を占める”という川村の原則に倣って(川村、1958)“メス”に重ね合わせれば、アイデンテイフィケーション・セオリーもいま少し命脈を保ち、他の研究者との議論も可能だったかもしれません。実を言うと、今西さんの文献の一部に「メスにも適用可能」とする記述が残っているのですが、それはあくまでも断片的言及にとどまります。こちらも、いまから振り返れば、メスにあまり関心を持たない一種のジェンダー・バイアスが働いていたのかもしれません。
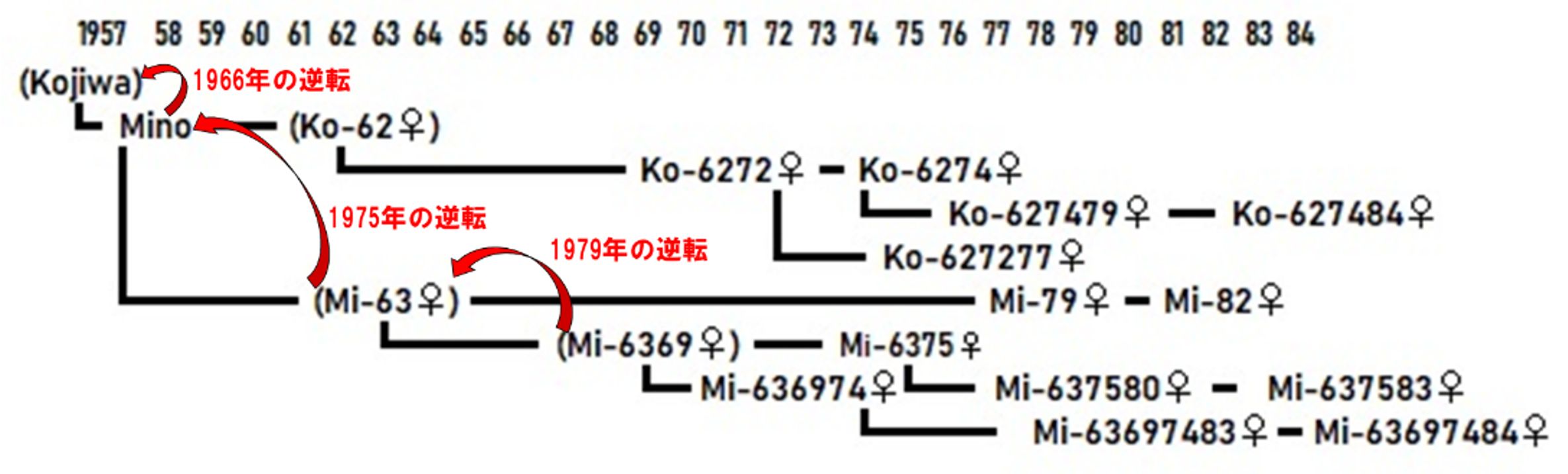
生まれた群れにとどまるニホンザルのメスたちは、その結果、母系的血縁を介し、数世代にもわたる血縁集団(家系)を形成します。上図は、嵐山での最優位血縁集団、コジワ(Kojiwa)を始祖とするKo家系における、1957〜84年に生まれたメスの血縁図です。もっとも、この家系ではきわめて例外的に、Kojiwa対Mino(1966年)、Mino対Mi-63♀(75年)、Mi-63♀対Mi-6369♀(79年)の3代にわたって、母親と長女の間で順位が逆転したため(図中の赤い矢印)、川村の法則に合致しない血縁関係の組合せが多数生じています。なお、これは“female competition”(メスの競争)における特異例として、詳細は省略します
伊谷先生は、今西さんとの論争が続いた当時を振り返って、回顧します。「母系社会論がお気に召さなかった今西先生とは、ずいぶん論争を重ねた。ゼミ中に私が座を立った間に、先生は「あの頑固なやつを何とかしてくれ」とおっしゃったというが、私は一歩も引かなかった。こういう血の出るような論敵が、先生以外にはなかったことを思うと、薄氷を踏んできた感がなくはない」『サル・ヒト・アフリカ』(76-77頁)。
それでは、チンパンジーはどうなのか? 伊谷先生はかなり長い間、決めかねていたようにも思われます。1972年の『霊長類の社会構造』の執筆時点では、チンパンジーの餌付け成功からまだ6年、資料は充分ではなく、「チンパンジーの単位集団は、母系でもなければ父系でもなく、非単系(non-unilineal)なのであり、いわば、よそからやってきた幾頭かのオスを中心にして結成されたアソシェーションのようなものだ」とやや曖昧に述べるにとどまります(『霊長類の社会構造』137頁)。
しかし、その5年後に出版された『チンパンジーの原野』では、どうやら“父系”と言ってもおかしくはなさそうだとして、単位集団は「成長した雄たちにとって終生を托するに足る器である」かもしれないと考えるとともに、「チンパンジーの単位集団は父系ではないかという川中(健二)君の仮定が実証される日が、私は近い将来にかならずやってくるという確信をつよめつつある」と記すに至ります(『チンパンジーの原野』150-151頁)。

左は出自集団にとどまるオスたち:「左から右に、カソンタ、カサンガ、カメマンフとカジャバラ」(Kグループ、1971年5月)。右はニホンザルのメスと違って、集団間を移籍するメスのチンパンジー:「K集団の若いメスのチャウシク(1971年5月)。のちにM集団に移籍、ライオンに食われ、最後をとげた」
伊谷先生がこうした社会構造論をさらにアップデートしたのが、1984年のトーマス・ハックスリー記念講演*でした。『霊長類の社会構造』から12年の歳月を経、野外からさらに多様な社会構造の存在があきらかになり、通時的構造による比較論を練り直すことを決意します。なによりも、この時点で、マハレのデータがさらに蓄積されたことで、チンパンジーの単位集団の父系的性格を確信できたことが大きな励みになったでしょう。(*:イギリス王立人類学研究所がトマス・ヘンリー・ハックスリーの業績をたたえるため、1900年から毎年、人類学に大きく貢献した人物に記念メダルを授与し、記念講演を開催します:これまでの主要な講演者は、文末のリストをご参照下さい)

左は『霊長類の社会構造』(1972)。中央はトーマス・ハックスリー記念講演を公刊した英語論文(“The evolution of primate social structures" Man [New Series] 20:593-611, 1985)。右は講演の日本語訳を収めた『霊長類社会の進化』(1987)
この講演で伊谷先生は、雌雄の移出入と集団の継承性という視点から、霊長類の基本的単位集団(BSU: Basic social unit)を以下の6タイプに分けました。12年で蓄積された資料は、霊長類の社会が思いのほか、複雑なことを示します。
まず、集団の継承性が保証されないタイプとして、次の3類型を指摘します。
1.単婚:両性のコドモが移出、移入がない(インドリ、ヨザル、テナガザル等)。なお、鳥類ではこの単婚(一夫一妻)による子育てが一般的です。
2.多夫一妻:複数のメスのうち繁殖するのは1頭だけで、メスのコドモの移出があります。その一方で、複数のオスが移出入します(クチヒゲタマリン等)
3.一夫多妻:オスの移入がなく、出自集団を移出したオスは他群出自のメスと単雄複雌的集団を形成。オスの子供が成長すると複雄複雌的構造に変化し、息子の継承の可能性があります(ゴリラ)。
次に、集団が次世代に継承されるタイプとして、以下の3類型をあげます。
4.双系:オス・メスともに移出入する構造で、伊谷先生は南米に生息するホエザル類と、アフリカに生息するアカコロブスをあげています。
5.母系:ニホンザルが典型で、オスのみが集団を移出入し、集団は母系で継承されます。
6.父系:母系と鏡像のように、メスのみが集団を移出し(出産すると、そのままそこの集団にとどまる)、集団はオスが継承します(チンパンジーとボノボ)。
この他、ゲラダヒヒとマントヒヒに重層構造が知られています。
それでは、この6つのタイプはどんな系統関係にあると考えられるでしょう? 下図は、伊谷先生が系統学的パラダイムにもとづいて、霊長類の社会構造の進化過程の復元を試みたものです。単婚から母系に至る道筋は南米に棲む広鼻猿類とオナガザル科を擬し、一夫多妻から父系に至る道筋に類人猿の系統をあてはめています。
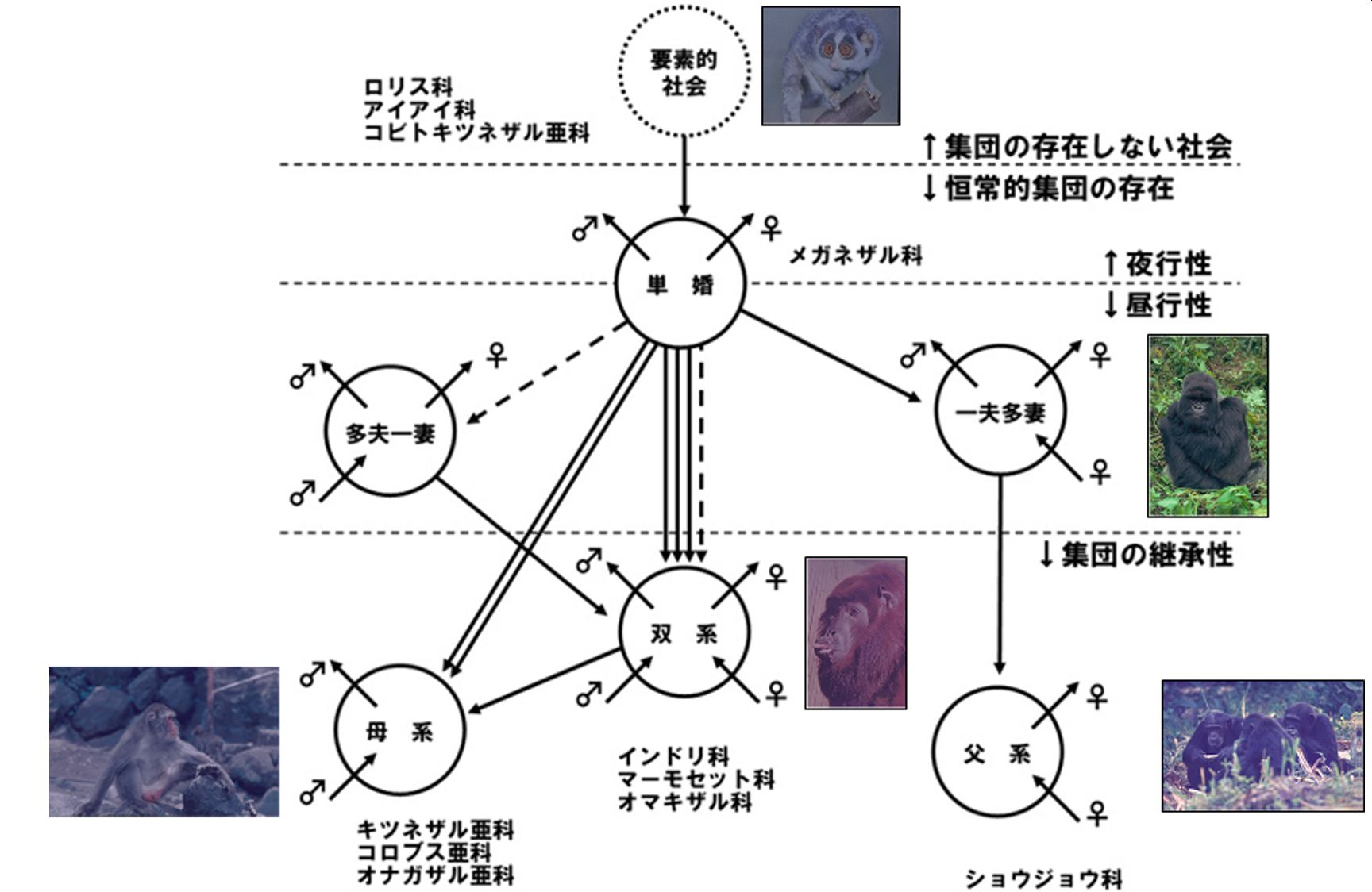
BSUをもたない要素的社会と、BSUの6型の関係。点線は推定
その上で、伊谷先生はこう述べます。「狭鼻猿類の二つの科は、オナガザル科が母系、ショウジョウ科が父系という実に対照的な分岐を見せるのである。この二つの構造は、その継承性の明確化からいっても、双系よりも完成度の高い構造と考えてよいだろう」「それでは、ヒトの社会はどの構造に由来するのであろうか。この問題は社会人類学の分野で多くの議論が交わされてきたが、私は、それが図の向かって右の系列に深い関係をもっているにちがいないこと、そしてチンパンジー属のBSUと狩猟採集民のバンド*の間には多くの類似性があるということだけを指摘しておきたいと思う」(『霊長類社会の進化』318-319頁)(*:採集狩猟民が血縁・婚姻関係等にもとづいて形成する、数十人から100人程度の小集団:第23回参照)
(以下、次号)
付録.主要なトーマス・ハックスリー記念講演者
1901年:フランシス・ゴルトン(イギリスの人類学・統計学・遺伝学者)
1903年:カール・ピアソン(イギリスの数理統計学・優生学者)
1914年:ジェームズ・フレイザー(イギリスの社会人類学・古典学者)
1922年:マルスラン・ブール(フランスの古生物学者;ネアンデルタール人研究)
1925年:サー・アーサー・ジョン・エヴァンズ(英国の考古学者;クノッソス遺跡発掘)
1936年:エドワード・ウェスターマーク(フィンランドの哲学・人類学者)
1938年:マルセル・モース(フランスの社会学・文化人類学者)
1944年:ヴィア・ゴードン・チャイルド(オーストラリア生まれの考古学・文献学者)
1945年:アルフレッド・ルイス・クローバー(米国の文化人類学者)
1950年:ジュリアン・ハクスリー(イギリスの進化生物学者、ヒューマニスト)
1951年:アルフレッド・ラドクリフ=ブラウン(イギリスの社会人類学者)
1956年:J・B・S・ホールデン(イギリスの生物学・集団遺伝学)
1963年:E・E・エヴァンズ=プリチャード(イギリスの社会人類学者)
1965年:C・レヴィ=ストロース(フランスの社会人類学者;構造主義人類学)
1971年:ジョージ・ピーター・マードック(米国の人類学者)
1980年:エドマンド・リーチ(イギリスの人類学者)
1984年:伊谷純一郎(日本の霊長類学者・生態人類学者)
1990年:ロバート・ハインド(イギリスの動物行動学者;行動生態学)
1992年:メアリー・ダグラス(英国社会人類・文化人類学者)
1998年:マーシャル・サーリンズ(アメリカの文化人類学者;文化進化論)
2000年:ピエール・ブルデュー(フランスの社会学・哲学者)
2002年:ジェーン・グドール(英国の霊長類・動物行動学者)
2004年:ティム・インゴルド(英国の社会人類学者)
2015年:ロビン・ダンバー(イギリスの生物学・人類学・進化生物学者)
編集・執筆:高畑由起夫
2025/12/5
一覧へ戻る